Вирусные болезни растений
Ви́рус (от лат. virus — яд) — микроскопическая частица, способная инфицировать клетки живых организмов. Вирусы являются облигатными паразитами — они не способны размножаться вне клетки. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружен также вирус, поражающий другие вирусы [1] . Вирусы представляют собой молекулы нуклеиновых кислот (ДНК или РНК), заключённые в защитную белковую оболочку (капсид). Наличие капсида отличает вирусы от других инфекционных агентов, вироидов. Вирусы содержат только один тип нуклеиновой кислоты: либо ДНК , либо РНК. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые белки и не содержат нуклеиновых кислот.
Положение вирусов в системе живого
Вирусы являются одной из самых распространённых форм существования органической материи на планете по численности: воды мирового океана содержат колоссальное количество бактериофагов (около 10 11 частиц на миллилитр воды).
Вирусы имеют генетические связи с представителями флоры и фауны Земли. Согласно последним исследованиям, геном человека более чем на 32 % состоит из информации, кодируемой вирус-подобными элементами и транспозонами. С помощью вирусов может происходить так называемый горизонтальный перенос генов (ксенология), то есть передача генетической информации не от отца к сыну и так далее, а между двумя неродственными (или даже относящимися к разным видам) особями. Так в геноме высших приматов существует белок синцитин, который, как считается, был привнесён ретровирусом. Иногда вирусы образуют с животными симбиоз [2] . Так, например, яд некоторых паразитических ос содержит структуры, называемые поли-ДНК-вирусами (Polydnavirus, PDV), имеющие вирусное происхождение.
Происхождение вирусов
Вирусы — сборная группа, не имеющая общего предка. В настоящее время существует несколько гипотез, объясняющих происхождение вирусов.
Считается, что крупные ядерно-цитоплазматические ДНК-содержащие вирусы происходят от более сложных (и, возможно, клеточных, таких как современные микоплазмы и риккетсии), внутриклеточных паразитов, утративших значительную часть своего генома. И действительно, некоторые крупные ДНК-содержащие вирусы (мимивирус, вирус оспы) кодируют функционально избыточные на первый взгляд ферменты, по-видимому, оставшиеся им в наследство от более сложных форм существования. Следует также отметить, что некоторые вирусные белки не обнаруживают никакой гомологии с белками бактерий, архей и эукариот, что свидетельствует о сравнительно давнем обособлении этой группы.
ДНК-содержащие бактериофаги и некоторые ДНК-содержащие вирусы эукариот, возможно происходят от мобильных элементов, участков ДНК способных к самостоятельной репликации в клетке.
Происхождение некоторых РНК-содержащих вирусов связывают с вироидами. Вироиды представляют собой высокоструктурированные кольцевые фрагменты РНК, реплицируемые клеточной РНК-полимеразой. Считается, что вироиды представляют собой «сбежавшие интроны» — вырезанные в ходе сплайсинга незначащие участки мРНК, которые случайно приобрели способность к репликации. Белков вироиды не кодируют. Считается, что приобретение вироидами кодирующих участков (открытой рамки считывания) и привело к появлению первых РНК-содержащих вирусов. И действительно, известны примеры вирусов, содержащих выраженные вироид-подобные участки (вирус гепатита Дельта).
Структура
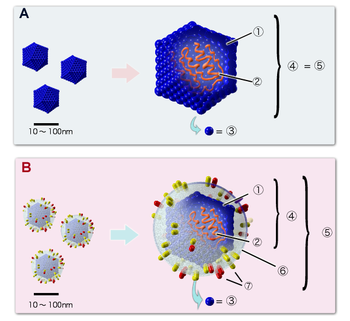
Примеры структур икосаэдрических вирионов.
А. Вирус, не имеющий липидной оболочки (например, пикорнавирус).
B. Оболочечный вирус (например, герпесвирус).
Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки.
Вирусные частицы (вирио́ны) представляют собой белковую капсулу — капсид, содержащую геном вируса, представленный одной или несколькими молекулами ДНК или РНК. Капсид построен из капсомеров — белковых комплексов, состоящих в свою очередь из протомеров. Нуклеиновая кислота в комплексе с белками обозначается термином нуклеокапсид. Некоторые вирусы имеют также внешнюю липидную оболочку. Размеры различных вирусов колеблются от 20 (пикорнавирусы) до 500 (мимивирусы) и более нанометров. Вирионы часто имеют правильную геометрическую форму (икосаэдр, цилиндр). Такая структура капсида предусматривает идентичность связей между составляющими её белками, и, следовательно, может быть построена из стандартных белков одного или нескольких видов, что позволяет вирусу экономить место в геноме.
Механизм инфицирования
Условно процесс вирусного инфицирования в масштабах одной клетки можно разбить на несколько взаимоперекрывающихся этапов:
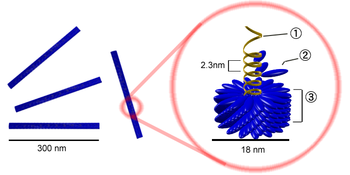
Палочковидная частица вируса табачной мозаики.
Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида.
- Присоединение к клеточной мембране — так называемая адсорбция. Обычно для того, чтобы вирион адсорбировался на поверхности клетки, она должна иметь в составе своей плазматической мембраны белок (часто гликопротеин) — рецептор, специфичный для данного вируса. Наличие рецептора нередко определяет круг хозяев данного вируса, а также его тканеспецифичность.
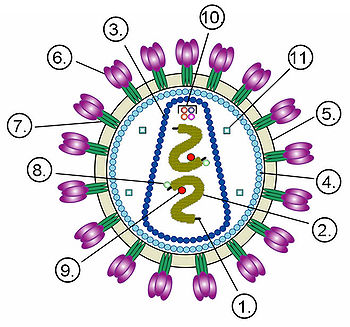
Структура вириона неикосаэдрического оболочечного вируса на примере ВИЧ.
Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — трансмембранный гликопротеин.
Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза.
- Проникновение в клетку. На следующем этапе вирусу необходимо доставить внутрь клетки свою генетическую информацию. Некоторые вирусы привносят также собственные белки, необходимые для её реализации (особенно это характерно для вирусов, содержащих негативные РНК). Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза, попадают в кислую среду лизосом, где происходит их окончательное созревание(депротеинизация вирусной частицы), после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает в цитоплазму. Вирусы также различаются по локализации их репликации, часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) в её ядре.
- Перепрограммирование клетки. При заражении вирусом в клетке активируются специальные механизмы противовирусной защиты. Заражённые клетки начинают синтезировать сигнальные молекулы — интерфероны, переводящие окружающие здоровые клетки в противовирусное состояние и активирующие системы иммунитета. Повреждения, вызываемые размножением вируса в клетке, могут быть обнаружены системами внутреннего клеточного контроля, и такая клетка должна будет «покончить жизнь самоубийством» в ходе процесса, называемого апоптозом или программируемой клеточной смерти. От способности вируса преодолевать системы противовирусной защиты напрямую зависит его выживание. Неудивительно, что многие вирусы (например, пикорнавирусы, флавивирусы) в ходе эволюции приобрели способность подавлять синтез интерферонов, апоптозную программу и так далее. Кроме подавления противовирусной защиты, вирусы стремятся создать в клетке максимально благоприятные условия для развития своего потомства. Хрестоматийным примером перепрограммирования систем клетки-хозяина является трансляцияРНКэнтеровирусов (семейство пикорнавирусы). Вирусная протеаза расщепляет клеточный белок eIF4G, необходимый для инициации трансляции подавляющего большинства клеточных мРНК (транслирующихся по так называемому кэп-зависимому механизму). При этом инициация трансляции РНК самого вируса происходит другим способом (IRES-зависимый механизм), для которого вполне достаточно отрезанного фрагмента eIF4G. Таким образом, вирусные РНК приобретают эксклюзивные «права» и не конкурируют за рибосомы с клеточными.
- Персистенция. Некоторые вирусы могут переходить в латентное состояние (так называемая персистенция для вирусов эукариот или лизогения для бактериофагов — вирусов бактерий), слабо вмешиваясь в процессы, происходящие в клетке, и активироваться лишь при определённых условиях. Так построена, например, стратегия размножения некоторых бактериофагов — до тех пор пока заражённая клетка находится в благоприятной среде, фаг не убивает её, наследуется дочерними клетками и нередко интегрируется в клеточный геном. Однако при попадании заражённой лизогенным фагом бактерии в неблагоприятную среду, возбудитель захватывает контроль над клеточными процессами так, что клетка начинает производить материалы, из которых строятся новые фаги (так называемая литическая стадия). Клетка превращается в фабрику, способную производить многие тысячи фагов. Зрелые частицы, выходя из клетки, разрывают клеточную мембрану, тем самым убивая клетку. С персистенцией вирусов (например, паповавирусов) связаны некоторые онкологическиезаболевания.
- Создание новых вирусных компонентов. Размножение вирусов в самом общем случае предусматривает три процесса — 1) транскрипция вирусного генома — то есть синтез вирусной мРНК, 2) её трансляция, то есть синтез вирусных белков и 3) репликация вирусного генома (в некоторых случаях, когда генетическая информация вируса закодирована в виде РНК геномная РНК одновременно играет роль мРНК, и, следовательно, процесс транскрипции в паразитируемой клетке не происходит за ненадобностью). У многих вирусов существуют системы контроля, обеспечивающие оптимальное расходование биоматериалов клетки-хозяина. Например, когда вирусной мРНК накоплено достаточно, транскрипция вирусного генома подавляется, а репликация напротив — активируется.
- Созревание вирионов и выход из клетки. В конце концов, новосинтезированные геномные РНК или ДНК одеваются соответсвующими белками и выходят из клетки. Следует сказать, что активно размножающийся вирус не всегда убивает клетку-хозяина. В некоторых случаях (например, ортомиксовирусы) дочерние вирусы отпочковываются от плазматической мембраны, не вызывая её разрыва. Таким образом, клетка может продолжать жить и продуцировать вирус.
Классификация
В таксономии живой природы вирусы выделяются в отдельный таксон Vira, образующий в классификации Systema Naturae 2000 вместе с доменами Bacteria, Archaea и Eukaryota корневой таксон Biota. В течение XX века в систематике выдвигались предложения о создании выделенного таксона для неклеточных форм жизни (Aphanobionta Novak, 1930; надцарство Acytota Jeffrey, 1971; Acellularia), однако такие предложения не были кодифицированы.
Систематику и таксономию вирусов кодифицирует и поддерживает Международный Комитет по Таксономии Вирусов (International Committee on Taxonomy of Viruses, ICTV), поддерживающий также и таксономическую базу The Universal Virus Database ICTVdB.
ICTV классификация
Международным Комитетом по Таксономии Вирусов в 1966 году была принята система классификации вирусов основанная на различии типа (РНК и ДНК), количества молекул нуклеотических кислот (одно- и двух-цепочечные) и на наличии или отсутствии оболочки ядра. Система классификации представляет собой серию иерархичных таксонов:
Отряд (-virales) Семейство (-viridae) Подсемейство (-virinae) Род (-virus) Вид (-virus)
Классификация Балтимора
Нобелевский лауреат, биолог Дэвид Балтимор, предложил свою схему классификации вирусов, основываясь на различиях в механизме продукции мРНК.Эта система включает в себя семь основных групп: [3] [4]
- (I) Вирусы, содержащие двуцепочечную ДНК и не имеющие РНК-стадии (например, герпесвирусы, поксвирусы, паповавирусы, мимивирус).
- (II) Вирусы, содержащие двуцепочечную РНК (например, ротавирусы).
- (III) Вирусы, содержащие одноцепочечную молекулу ДНК (например, парвовирусы).
- (IV) Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности (например, пикорнавирусы, флавивирусы).
- (V) Вирусы, содержащие одноцепочечную молекулу РНК негативной или двойной полярности (например, ортомиксовирусы, филовирусы).
- (VI) Вирусы, содержащие одноцепочечную молекулу РНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретровирусы (например, ВИЧ).
- (VII) Вирусы, содержащие двуцепочечную ДНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретроидные вирусы (например, вирус гепатита B).
В настоящее время, для классификации вирусов используются обе системы одновременно, как дополняющие друг друга. [5] [6] [7]
Дальнейшее деление производится на основе таких признаков как структура генома (наличие сегментов, кольцевая или линейная молекула), генетическое сходство с другими вирусами, наличие липидной оболочки, таксономическая принадлежнось организма-хозяина и так далее.
История
В 2002 в университете Нью-Йорка был создан первый синтетический вирус (вирус полиомиелита).
Применение вирусов
- Генная инженерия
- Нанотехнологии
- Биологическое оружие
Источник https://dic.academic.ru/dic.nsf/ruwiki/844259